


Augenarzt in München
Die Augenarztpraxis Prof. Dr. Marcus Kernt & Kollegen
Prof. Dr. Marcus Kernt
Nach Abschluss seines Medizinstudiums an der Ludwig-Maximilians-Universität (LMU) München und der Assistenzarztzeit an der Augenklinik der LMU erwarb Augenarzt Prof. Dr. Kernt die Anerkennung als Facharzt für Augenheilkunde. Bereits während seiner Ausbildungszeit bei Herrn Professor Anselm Kampik an der Universitätsaugenklinik in München gewann er umfassende Kenntnisse in allen Bereichen der Augenheilkunde mit besonderem Schwerpunkt auf Erkrankungen der Netzhaut und der refraktiven Chirurgie.
Mit dem Erwerben der Facharztreife war Prof. Dr. Kernt viele Jahre oberärztlich an der Augenklinik der LMU tätig und ist seit Januar 2014 auch als Augenarzt in München-Fürstenried niedergelassen. Neben seiner umfangreichen klinischen und wissenschaftlichen Tätigkeit absolvierte er mehrere Forschungs- und Weiterbildungsaufenthalte im Ausland. Prof. Kernt war im Moorefields Eye Hospital in London tätig, wo er sich neben Retinologie und Vitreoretinal-Chirurgie bei Prof Alan Bird und Zdenek J. Gregor auch intensiv in Okuloplastik und Lid-Chirurgie bei Prof. Richard Collin, einem der weltweit führendsten Spezialisten auf diesem Gebiet, weiterbildete. Aus den hieraus entstanden Kontakten haben sich über die Jahre wissenschaftliche Kooperationen mit englischen Institutionen ergeben, so auch mit der Universität Oxford, wo Augenarzt Prof. Kernt seit 2015 auch reguläres Mitglied des Lehrkörpers ist.

Mehr zu Prof. Dr. M. Kernt
2003 bis 2004 arbeitete er unter der Leitung von Prof. Alain Gaudric am Hôpital Lariboisiere in Paris und 2013 im Narayana Eye Hospital in Bangalore auf dem Gebiet der Glaskörperchirurgie. Im Rahmen seiner oberärztlichen Tätigkeit an der Universitätsaugenklinik München leitete er über mehrere Jahre zusammen mit Prof. Ulbig und Prof. Haritoglou die Spezialsprechstunden für Diabetische Augenerkrankungen und Altersbedingte Makuladegeneration. Als stellvertretender Leiter des an der LMU ansässigen ‚Centrum für Refraktive Therapie CRT‘ arbeitete er auch im Bereich der refraktiven Chirurgie.
Im Januar 2011 habilitierte Augenarzt Prof. Dr. Kernt an der Ludwig-Maximilians-Universität München im Fach Augenheilkunde. Seine wissenschaftliche Arbeit umfasst neben der zellbiologischen Forschung vor allem die klinische Erforschung neuer Behandlungsmethoden im Bereich vaskulärer Netzhauterkrankungen und des Glaukoms, aber auch die ophthalmologische Bildgebung und Diagnostik.
2015 erhielt Prof. Kernt einen Ruf der englischen Elite-Universität Oxford und wurde dort im September dieses Jahres zum ordentlichen Professor für Ophthalmochirurgie und Health Care Management ernannt. Im Rahmen seiner akademischen Forschungstätigkeit hat Augenarzt Prof. Dr. Kernt an einer Vielzahl von wissenschaftlichen Veröffentlichungen mitgewirkt, zahlreiche Vorträge auf nationalen und internationalen Kongressen gehalten und wurde mit eine Reihe hochrangiger wissenschaftlicher Förderungen, Preise und Ehrungen bedacht. So wurde Augenarzt Prof. Dr. Kernt im Jahr 2013 für seine herausragende wissenschaftliche Arbeit im deutschsprachigen Raum mit dem namhaften Theodor-Axenfeld-Preis ausgezeichnet. Herr Prof. Dr. Kernt verfügt über die volle Lehrbefugnis im Bereich Augenheilkunde, außerdem hält Augenarzt Prof. Dr. Kernt eine reguläre Professorenstelle an der Universität Oxford inne und ist dort ebenfalls reguläres Mitglied des Lehrkörpers. Seit Januar 2014 ist Augenarzt Prof. Dr. Kernt neben seiner akademischen Tätigkeit schwerpunktmäßig als Augenarzt in seinen Facharztpraxen in München-Fürstenried und Grünwald tätig.
Werdegang
Studium
- Christian-Albrechts-Universität, Kiel
- Ludwig-Maximilians-Universität, München
Ophthalmologische Weiterbildung und akademischer Werdegang
- Wissenschaftlicher Weiterbildungsassistent an der Augenklinik, Klinikum der LMU München (2004-2010)
- Facharzt für Augenheilkunde an der Augenklinik, Klinikum der LMU München (2010)
- Habilitation im Fach Augenheilkunde an der Medizinischen Fakultät der LMU München (2011)
- Berufung zum ordentlichen Universitätsprofessor, University of Oxford, Oxford, UK (2015)
Auslandsaufenthalte
- Moorefields Eye Hospital, London (bei Prof. A. Bird, Z.J. Gregor und J.R.O. Collin, 2002)
- Hôpital Lariboisiere, Paris (bei Prof. A. Gaudric, 2003-2004)
- Narayana Eye Hospital, Bangalore (bei Dr Rohit Shetty)
Klinische Schwerpunkte
- Erkrankungen der Netzhaut und des Glaskörpers mit Schwerpunkt auf Diabetischen Augenerkrankungen (Diabetische Retinopathie, Diabetisches Makulaödem) und Altersabhängige Makuladegeneration
- Erkrankungen der Hornhaut und des vorderen Augenabschnitts, Katarakt (Grauer Star)
- Glaukom
Operatives Spektrum
- Behandlung des Grauen Stars (Katarakt-Chirurgie)
- Operative Behandlung von Fehlsichtigkeiten (Excimer-Laser-Chirurgie: Lasik, PRK; Intraokularlinsen-Chirurgie)
- Glaskörperchirurgie
- Glaukom-Laserchirurgie
- Netzhaut-Lasertherapie
Forschungsschwerpunkte
- Altersbedingte Makuladegeneration
- Diabetische Retinopathie
- Ophthalmologische Bildgebung
- Zellbiologische Forschung
Mitgliedschaften in Fachgesellschaften
- American Academy of Ophthalmology (AAO)
- Association for Research in Vision and Ophthalmology (ARVO)
- Berufsverband der Augenärzte Deutschlands e.V. (BVA)
- Bundesverband Deutscher Ophthalmochirurgen (BDOC)
- Deutsche Ophthalmologische Gesellschaft (DOG)
- European Association for Vison and Eye Research (EVER)
- European Society of Cataract and Refractive Surgeons (ESCRS)
- European Society of Retina Specialists (EURETINA)
- Société Française d’Ophtalmologie (SFO)
- Prof. Dr. Kernt ist zertifizierter Anwender der Kommission für Refraktive Chirurgie (KRC)
Mitwirkung bei internationalen Fachjournalen
- „Acta Ophthalmologica“
Reviewer für:
- “American Journal of Ophthalmology”
- “British Journal of Ophthalmology”
- “Cornea”
- “Diabetes Care”
- “Eye”
- “Graefes Archives of Ophthalmology”
- “Indian Journal of Ophthalmology”
- “JAMA Ophthalmology”
- “Journal of Biomatherial Science”
- “Journal of Cataract and Refractive Surgery”
- “Journal of Clinical Ophthalmology”
- “Toxicology”
Unsere Augenarztpraxis in München
Gutes Sehen bedeutet Freiheit, Selbstbestimmung, Unabhängigkeit, sowohl im Beruf aber auch in der Freizeit. Der Mensch nimmt seine Umwelt zu über 80% durch die Augen wahr. Ein gutes Sehvermögen trägt ganz entscheidend zur Lebensqualität bei. Ist das Sehvermögen beeinträchtigt, stellen wir sehr schnell fest, wie elementar wichtig gutes Sehen im Alltag ist. Um Ihr gutes Sehen zu erhalten oder aber auch wieder zu verbessern, stellt der Augenarzt in München Prof. Dr. Kernt & Kollegen Ihnen ein Höchstmaß an fachlicher Erfahrung, Kompetenz, aber auch modernste diagnostische und therapeutische Möglichkeiten zur Verfügung.
Ihr gutes Sehen ist unser Auftrag
Wir möchten Ihnen die beste augenärztliche Medizin anbieten und können hierbei auf ein breites Spektrum an hochmodernen Diagnostik- und Behandlungsmöglichkeiten zurückgreifen. Diese Internetseite soll Ihnen einen Überblick über unsere Leistungen bieten und Ihnen zudem die Möglichkeit geben, sich über unsere Praxis und deren medizinischem Angebot zu informieren, mit dem wir Ihnen, aus ganzem Herzen und mit vollem Einsatz, zur Seite stehen wollen. Augenarzt in München Dr. Kernt und sein Team freuen sich, Ihnen zu einem guten Sehen verhelfen zu können!
Um Ihnen die bestmögliche Behandlung zukommen zu lassen, setzen wir sowohl bei Diagnostik, als auch Therapie auf modernste Technologie und die höchsten medizinischen Standards. Dabei können Sie sich auf die hervorragende fachliche Kompetenz und Erfahrung von Augenarzt Prof. Dr. med. habil. Marcus Kernt und seinen Kollegen im Bereich der Augenheilkunde verlassen. Die apparativ-instrumentelle Ausstattung unserer Praxis ermöglicht es uns Ihnen Augenheilkunde auf höchsten wissenschaftlichen Niveau und mit den modernsten technischen Möglichkeiten anzubieten, die denen einer Augenklinik vergleichbar sind. Dennoch legen wir auch besonderen Wert auf individuelle Beratung und Betreuung, damit Sie sich wohl fühlen und Ihre Beschwerden bestmöglich behandelt werden! Dank der guten Verkehrsanbindung (U-Bahn und Bus direkt vor der Praxistür – Haltestelle „Forstenrieder Allee“ -, sowie zahlreichen Parkmöglichkeiten in der Nähe und des behindertengerechten Ausbaus, erreichen Sie uns leicht und mühelos im Herzen von München-Fürstenried.
Augenarzt Prof. Dr. Kernt trägt außerdem regelmäßig mit seinem Fachwissen dazu bei, Patienten über das medizinische Portal info Medizin mit Informationen rund um die Augenheilkunde zu versorgen.
Ihr Augenarzt in München
Prof. Dr. Marcus Kernt
Impressionen
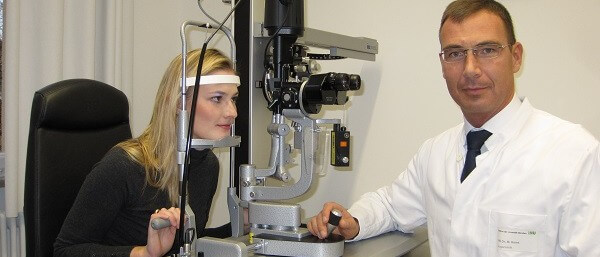




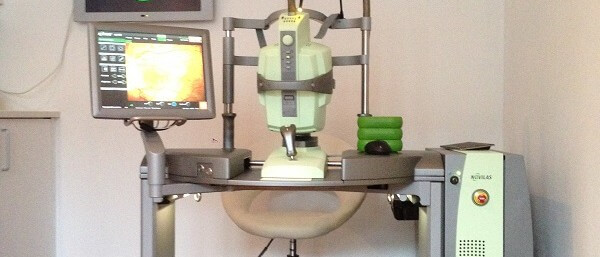
Forschung und Wissenschaft sind die Grundlage medizinischen Fortschritts
Forschung und Wissenschaft sind die Grundlage medizinischen Fortschritts! Nur wer weiter denkt, kann auch in der Medizin und Augenheilkunde die Behandlungsoptionen für seine Patienten erweitern und verbessern. Evidenzbasierte Medizin ist eine wichtige Voraussetzung für patientenorientierte Entscheidungen auf der Grundlage von empirisch nachgewiesener Wirksamkeit. Augenarzt Prof. Dr. med. habil. Marcus Kernt hat sich über viele Jahre intensiv, sowohl mit klinischer Forschung, aber auch mit ophthalmologischer Grundlagenforschung beschäftigt und sich hierbei national und international einen Namen gemacht.
Neben seiner Tätigkeit als niedergelassener Augenarzt in München ist Herr Prof. Dr. Kernt Mitglied des Lehrkörpers der Ludwig-Maximilians-Universität München und geht hierbei einer regelmäßigen Lehrtätigkeit nach. Er wirkt an einer Reihe klinischer Studien mit und hat neben einer Vielzahl von Kongressbeiträgen auch an mehr als hundert wissenschaftlichen Veröffentlichungen mitgewirkt. Auf dieser Seite finden Sie eine Auswahl der von Prof. Dr. Kernt veröffentlichten Fachartikeln aus den unterschiedlichsten Teilgebieten der Augenheilkunde. Bei Interesse informieren wir Sie gerne über die aktuelle wissenschaftliche Arbeit von Augenarzt Prof. Dr. Kernt und wie diese hilft Ihre Behandlung zu verbessern.
Fachartikel von Prof. Dr. Marcus Kernt
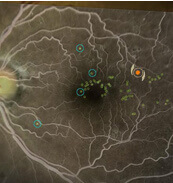
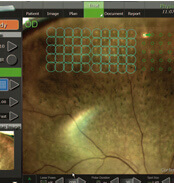
Navigierte Netzhautlasertherapie
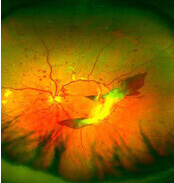
Diabetische Retinopathie
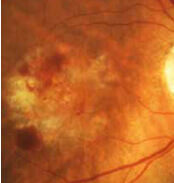
Neue therapeutische Möglichkeiten
Bilddiagnostik und Lasertherapie kombiniert
Diabetisches Makulaödem
Anti-VEGF-Therapien bei der Makuladegeneration

Protektive Wirkung Blaulicht absorbierender IOLs auf Pigmentepithel
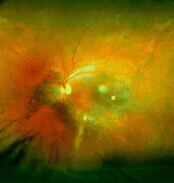
Weitwinkel- Fundusautofluoreszenz bei Uveitis posterior
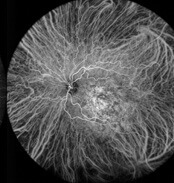
Differenzierung pigmentierter Fundusveränderungen
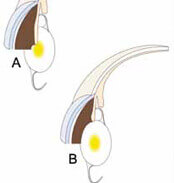
Ein neues IOL-Konzept für optimalen Netzhautschutz?
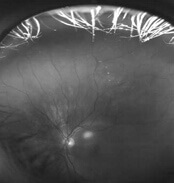
Imaging peripherer Netzhautveränderungen
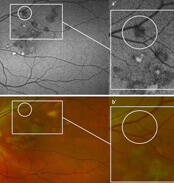
Weitwinkel – fundusautofluoreszenz bei Uveitis posterior
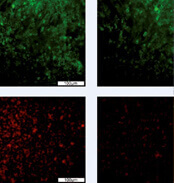
Zytoprotektive Wirkung des Multikinaseinhibitors
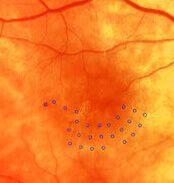
Navigierte fokale retinale Lasertherapie mit dem NAVILAS®-System
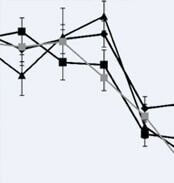
Moxifloxacin intrakameral

Verbesserung des Fixationsverhaltens bei diabetischem Makulaödem

Multikinase-Inhibitoren bei neovaskulärer AMD
Unsere Leistungen
Finden Sie hier eine Übersicht über unsere augenärztlichen Leistungen in München:
Therapie
- Hornhaut-Laserbehandlung (Refraktive Hornhaut-Laserchirurgie)
- Femto-Lasik
- PRK (Photorefraktive Keratektomie)
- Behandlung des grünen Stars
- SLT (Selektive Lasertrabekuloplastik)
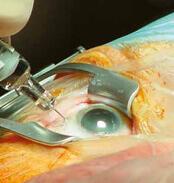
- Behandlung des grauen Stars
- Monofokale Intraokularlinsen
- Premium Intraokularlinsen
- Asphärische Intraokularlinsen
- Torische Intraokularlinsen
- Multifokale Intraokularlinsen
- Pseudoakkommodative Intraokularlinsen
- „Gelbe“ Blaulichtfilterlinsen
- Nachstarbehandlung
- Refraktive Linsenchirurgie
- Netzhaut-Laserbehandlung
- Navigierte Netzhaut-Lasertherapie (NAVILAS®)
- NAVILAS® Prime Laser System
- IVOM (Medikamenteneingabe in den Glaskörperraum)
- Glaskörperchirurgie
Diagnostik
- 3D-Katarakt-Analyse
- Pachymetrie (Hornhautdickenmessung)
- Vorderkammeranalyse
- Optische Biometrie des Auges
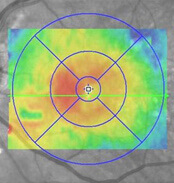
- Kohärenztomographie (OCT)
- Netzhautuntersuchung mit OPTOMAP®
- Fluoreszenzangiographie
- Autofluoreszenz
- Fundusfotographie
Aktuelles
Kontakt
Augenarzt
Prof. Marcus Kernt & Kollegen
Forstenrieder Allee 59
81476 München
Sprechzeiten
Mo - Mi. 9:00 - 18:00 Uhr
Do. nach Vereinbarung
Fr. 9:00 - 15:00
sowie nach Vereinbarung
Terminvereinbarung
Wir freuen uns Sie in unserer Augenarztpraxis in München begrüßen zu dürfen. Zur Terminvereinbarung rufen Sie an oder schreiben Sie uns über unser Kontaktformular:
